


L’Ordovicien est le second système du Phanérozoïque, l’éon qui a vu le développement du règne animal. Succédant au Cambrien, il a commencé il y a 485 millions d’années et a duré 42 millions d’années. C’est le géologue britannique Charles Lapworth qui l’a défini en 1879. Son nom a été emprunté à la tribu celtique des Ordovices, du pays de Galles. La terminologie anglaise s’applique toujours à deux subdivisions de l’Ordovicien : le Trémadocien (de 485 à 478 Ma), à sa base, doit son nom à Tremadoc au nord du Pays de Galles, et le Hirnantien (de 445 à 443 Ma), à son sommet, a été défini à la localité de Cwm Hirnant près de Bala, dans la même région. Elle continue à être utilisée de manière non officielle pour d’autres subdivisions, comme le Caradoc et l’Ashgill, constituant l’Ordovicien supérieur (de 458 à 443 Ma).
La paléogéographie de l’Ordovicien
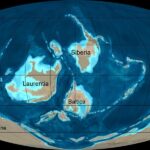
Les continents actuels de l’hémisphère Sud étaient regroupés au sein du Gondwana, avec l’Australie et le craton de Chine du Nord en partie situés au nord de l’équateur. Celui-ci traversait la Siberia, alors isolée dans l’océan mondial, et la Laurentia, comprenant l’Amérique du Nord et le Groenland, ainsi que le nord-ouest de l’Irlande et de l’Écosse. La Baltica, le noyau de l’Europe, se trouvait dans l’hémisphère Sud. Il était séparé de la Laurentia par l’océan Iapetus, dont la fermeture a commencé durant le Cambrien. Elle a entraîné la surrection de plusieurs montagnes, série d’évènements qui est appelée l’orogenèse calédonienne (ou le cycle orogénique calédonien) et qui a duré jusqu’au début du Dévonien.
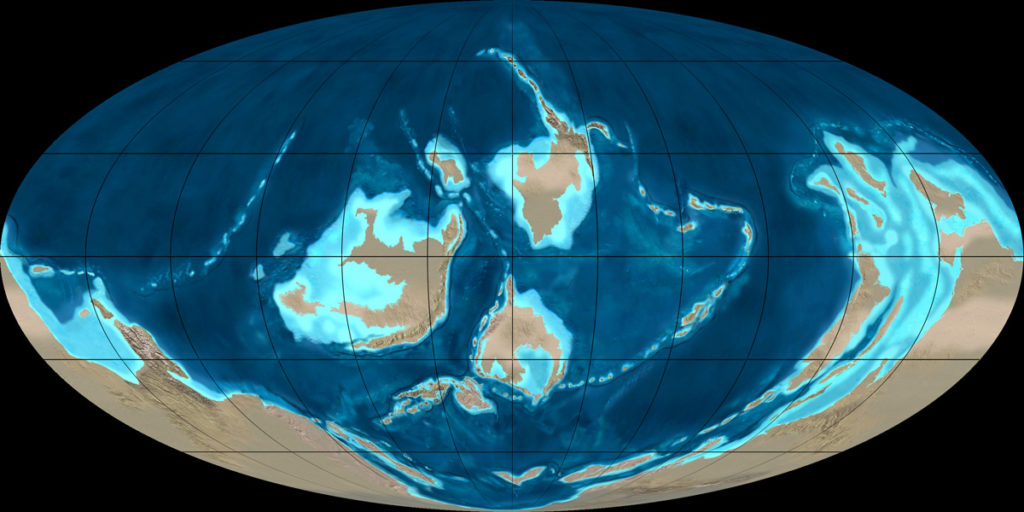
Au cours du Cambrien, la subduction de la plaque de la Laurentia, en partie océanique, sous la plaque du Iapetus, a fait naître un arc d’îles volcaniques. Ce microcontinent est entré en collision avec la Laurentia du côté de l’Irlande et de l’Écosse pendant l’Ordovicien inférieur et moyen (475-460 Ma), créant la phase grampienne de cette orogenèse. Le cycle s’est poursuivi à la fin de l’Ordovicien avec la phase taconienne, dont le nom provient des monts Tacon dans l’État de New York. Elle est due à la collision de l’arc avec la partie nord-américaine de la Laurentia.
Ailleurs dans le monde, l’Ordovicien a été une période de transgressions, c’est-à-dire d’avancée des mers sur les continents. Le phénomène a commencé durant le Cambrien et s’est poursuivi au cours de l’Ordovicien inférieur. Il y a eu une régression à la fin de l’Ordovicien moyen, qui correspond à un refroidissement et une radiation évolutive décrits ci-dessous, puis le niveau des mers a de nouveau monté. Il a atteint son maximum pendant l’Ordovicien supérieur et a baissé à cause de la grande glaciation hirnantienne. Puisque de grandes mers épicontinentales se sont constituées, les roches de l’Ordovicien sont surtout sédimentaires et elles sont dominées par les carbonates : calcaires et dolomies. On en trouve au sommet de l’Everest, dans la formation dite de Qomolangma. Mais au Pays de Galles, la sédimentation a été détritique : il s’agit surtout de grès. Il y a également des laves basaltiques et de la rhyolite.
La faune de l’Ordovicien
Ces transgressions ont été profitables à la vie, qui était encore à peu près cantonnée dans les mers et les océans. Comme durant le Cambrien, le règne animal était encore très largement dominé par les invertébrés. Les embranchements actuels existaient déjà : les annélides, les arthropodes, les échinodermes, les brachiopodes, les mollusques, etc. Les brachiopodes, peu représentés actuellement, sont des animaux à coquille composée de deux valves, mais la symétrie n’est pas la même que chez les mollusques bivalves. Ils ont cependant la symétrie bilatérale en commun. Les brachiopodes sont des protostomiens (l’un des deux groupes de bilatériens), de même que les bryozoaires, les « animaux mousses » apparus au cours de l’Ordovicien inférieur, vivant en colonies semblables à des éventails de dentelle et parfois confondus avec des coraux. Chaque individu sécrète un squelette calcaire relié à celui de ses voisins.
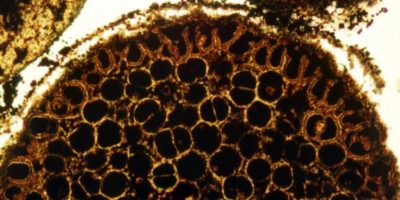
De nombreux animaux de l’Ordovicien, éteints depuis des centaines de millions d’années, nous semblent aujourd’hui très exotiques. C’est le cas des graptolites. Ils appartenaient à l’embranchement des hémichordés, proche des embranchements des échinodermes et des chordés (tous étant des deutérostomiens, l’autre groupe de bilatériens). Ils se répartissaient en deux groupes principaux : les dendroïdes et les graptoloïdes. Ils formaient tous des colonies appelées des rhabdosomes. Apparus au cours du Cambrien, il y a 520 Ma, les dendroïdes étaient des animaux benthiques et sessiles, c’est-à-dire qu’ils vivaient fixés sur le fond des mers. La fin du Cambrien voit l’apparition des graptoloïdes, qui étaient également coloniaux mais avaient un mode de vie planctonique : ils dérivaient sous la surface de l’eau. Les fossiles conservés provenaient de tubes protéïques protégeant ces animaux à corps mou. Longtemps difficiles à identifier, les graptolites ont été rapprochés dans les années 1990 des ptérobranches, des sortes de vers marins vivant toujours actuellement, au sein des hémichordés. Parmi les graptolites de l’Ordovicien inférieur, on peut citer Dendrograptus, retrouvé dans les sédiments de la Scandinavie et de l’Europe de l’Est. Dicanograptus vivait dans les océans de l’hémisphère Nord il y a 450 Ma.

Les crinoïdes ou lys de mer sont des échinodermes ressemblant à des plantes, apparus durant l’Ordovicien inférieur et abondants dans le registre fossile. À l’âge adulte, ils vivent fixés sur le fond marin, certains à de grandes profondeurs. La première espèce identifiée, appelée Athenacrinus broweni, est datée à 480 Ma. Les crinoïdes existent toujours, contrairement aux blastoïdes, apparus durant le Cambrien, en déclin à la fin du Carbonifère et disparus durant la crise de la fin du Permien, et aux cystoïdes, éteints durant le Dévonien.
Les mollusques regroupent les bivalves, apparus durant le Cambrien mais diversifiés à partir de l’Ordovicien inférieur, et les scaphopodes, des animaux pourvus de tubes de calcaire, apparus il y a 460 Ma durant l’Ordovicien moyen et toujours présents. Ils comprennent également les viscéroconques, divisés en gastéropodes et en céphalopodes (pieuvres, calmars, sèches, ainsi qu’ammonoïdes jusqu’au Crétacé et nautiloïdes). Certains de ces animaux sont devenus des monstres des mers, comme Cameroceras, un nautiloïde de 6 mètres de long. Il appartenait, comme Endoceras, dont les dimensions étaient semblables, à l’ordre des Endocéridés. À cause d’eux, l’Ordovicien est considéré comme « l’âge des nautiloïdes ». Ils se nourrissaient de trilobites et d’euryptérides, surnommés les scorpions de mer. C’était des arthropodes. L’un des premiers euryptérides connus, Megalograptus, a vécu durant l’Ordovicien supérieur. Ils sont devenus les super-prédateurs des mers de l’Ashgill et du Silurien. Les limules, proches d’eux, sont apparus un peu plus tard et vivent toujours dans les océans. Ce sont des chélicérates, de même que les arachnides (araignées et scorpions).

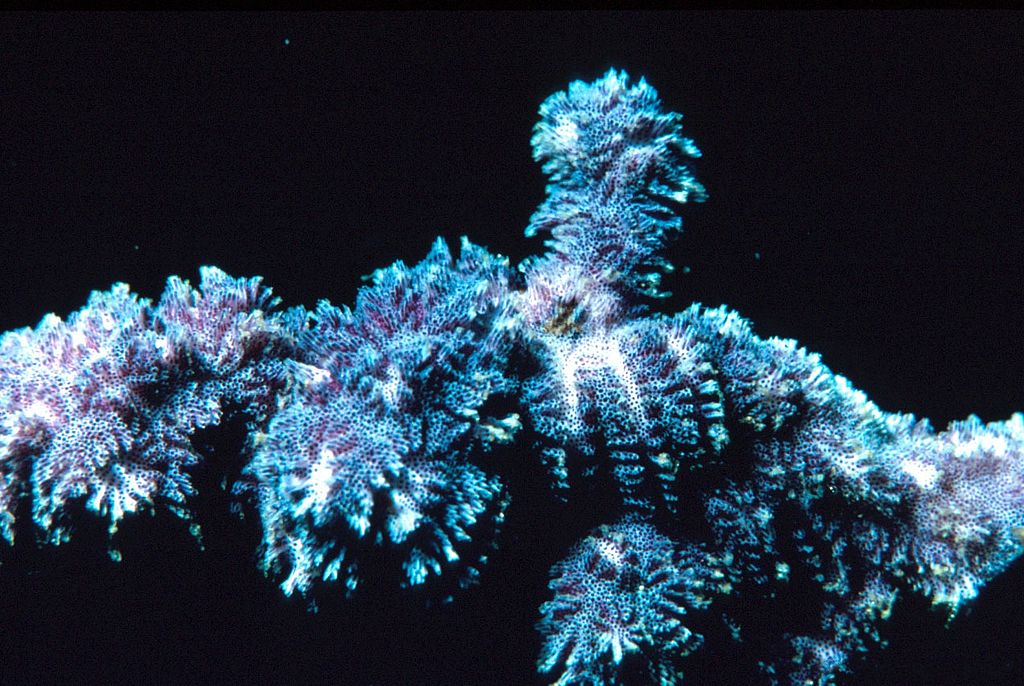
Si les actuels constructeurs de récifs sont les coraux, ce n’était pas le cas durant l’Ordovicien. Les premiers coraux identifiés datent d’il y a 450 Ma. Ce sont des coraux tabulés. Leur nom provient de la tabula, une cloison en calcaire séparant les polypes. Les coraux rugueux viennent ensuite, mais ils ne formeront de grands récifs qu’au cours du Silurien. Durant l’Ordovicien inférieur, les récifs étaient construits par des éponges, des organismes appelés des Calathium qui sont longtemps restés énigmatiques, ainsi que des bryozoaires à partir de la fin du Trémadocien. Il s’est avéré que ces Calathium étaient des éponges plutôt que des algues. Celles-ci ont commencé à jouer un rôle à partir de l’Ordovicien moyen, avec les stromatophores, qui étaient des éponges à squelette massivement calcifié. Ainsi, l’Ordovicien apparaît comme une importante période de transition.

Les vertébrés font partie de l’embranchement des chordés (ou cordés) mentionné ci-dessus. Avec eux, il y a aussi les céphalocordés, dont le représentant le plus connu est l’amphioxus. Ce petit animal sans squelette ni cerveau donne une idée de ce qu’étaient les ancêtres des vertébrés. Ceux-ci sont définis comme des chordés dotés d’un crâne permettant la protection du cerveau. Les tout premiers vertébrés sont appelés des agnathes car ils n’avaient pas encore de mâchoire, ce qui est encore le cas de la lamproie. Le premier agnathe, ou « poisson sans mâchoire », connu est Arandaspis, et date d’il y a 480 Ma. Il a été trouvé dans l’actuelle Australie. Sacabambaspis apparaît 10 millions d’années plus tard dans l’actuelle Bolivie. Ils étaient tous les deux gondwaniens. Deux autres agnathes, Astraspis et Eriptychius, ont vécu autour de la Laurentia et de la Baltica durant l’Ordovicien moyen.
Radiation évolution et extinction de masse
À partir l’Ordovicien moyen, plus particulièrement durant le Darriwilien (de 467 à 458 Ma), une grande diversification de la faune se produit. Il n’apparaît pas de nouveaux embranchements, mais le nombre de familles est quadruplé et le nombre de genres est triplé. Cette évolution se fait au détriment de la faune héritée du Cambrien, dominée par les arthropodes. Les mers acquièrent l’aspect qu’elles garderont durant le Paléozoïque, avec un plus grand nombre d’animaux pélagiques.
Plusieurs théories ont été proposées pour expliquer cette diversification. Les transgressions sur des continents dispersés ont probablement joué. En 2008, une équipe de scientifiques conduite par Julie Trotter a fourni une estimation des températures de surface des mers basée sur l’analyse des conodontes, de minuscules fossiles de forme de dents très utilisés en stratigraphie. Il s’avère que des mers ordoviciennes étaient très chaudes au début, avec des températures dépassant les 40 °C. Elles ont baissé jusqu’au Darriwilien, au moment de la grande diversification. Ces mers étaient alors comparables aux mers équatoriales actuelles : il y fait environ 30 °C. Elles étaient ainsi plus favorables à la vie. D’autres scientifiques ont expliqué ce refroidissement par les abondantes chutes de météorites qui ont été observées à cette époque.
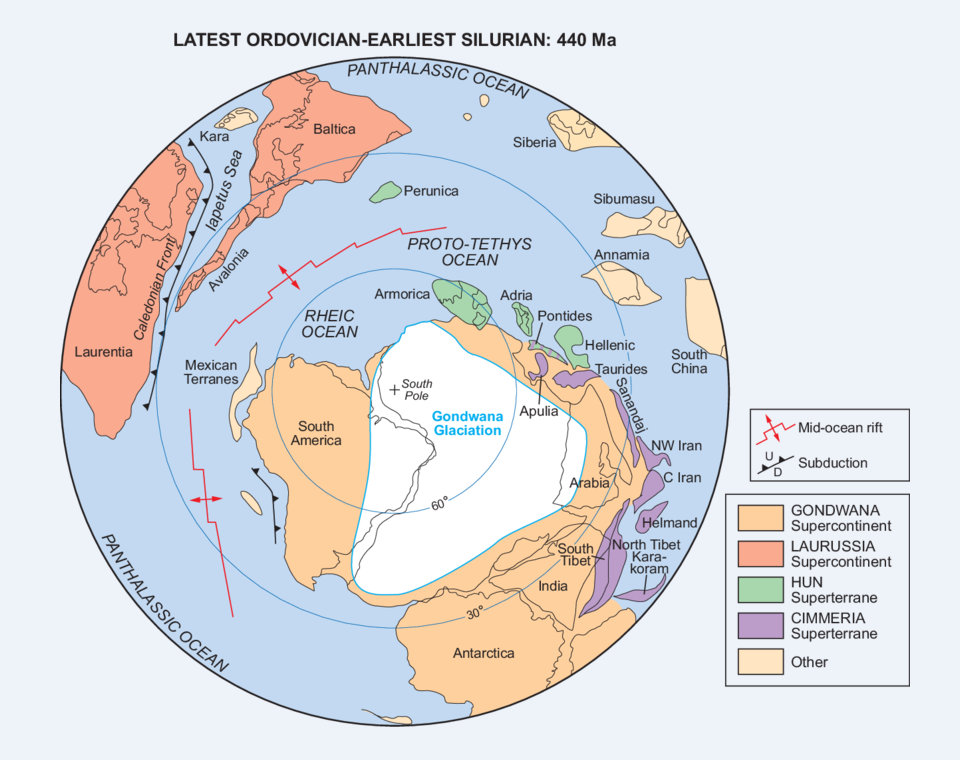
L’Ordovicien s’achève par la première extinction de masse du Phanérozoïque. La moitié des genres et 85% des espèces marines disparaissent. Elle coïncide avec la glaciation de l’Hirnantien. Ses traces sont abondantes dans le Sahara, où l’on trouve d’anciennes moraines et des roches striées par l’écoulement de glaciers. Elles sont également présentes dans la péninsule arabique. Ces traces n’existent pas en Afrique centrale, simplement parce qu’elles n’ont pas été conservées, mais on les retrouve en Afrique du Sud. Elles sont beaucoup moins répandues en Amérique du Sud. Il y a des témoignages de glaciation ordovicienne en Argentine occidentale et en Bolivie, et peut-être au Brésil.
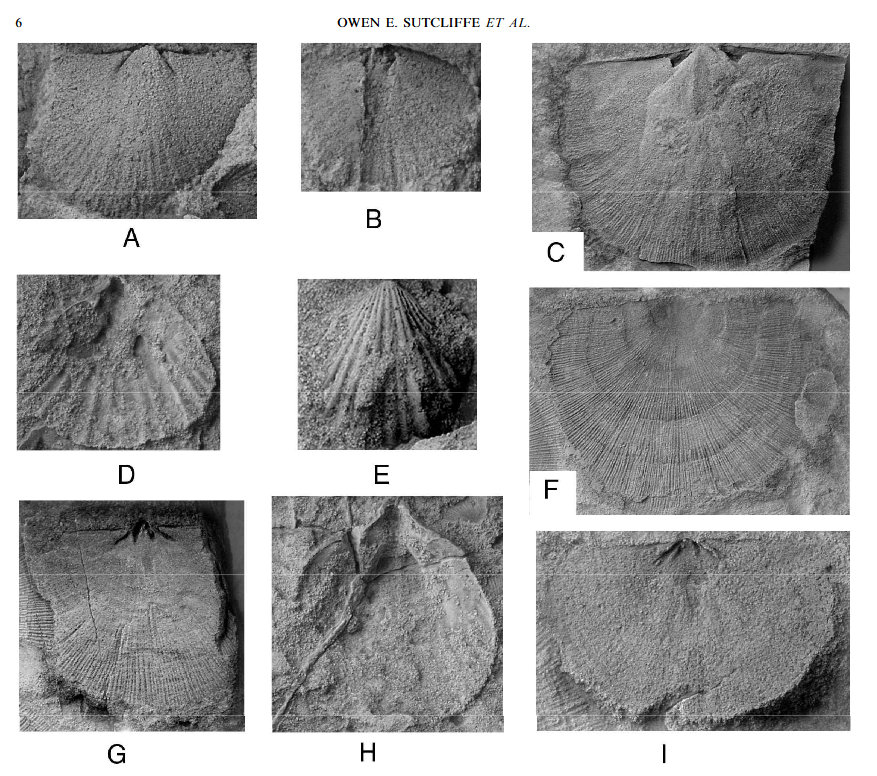
Comme l’Afrique et l’Amérique étaient voisines au sein du supercontinent Gondwana, cela implique l’existence d’une grande calotte glaciaire. Durant l’Hirnantien, le pôle Sud se trouvait dans l’actuel Sahara occidental. L’aire de cette calotte a été évaluée à 30 000 000 km², moins que celle des calottes du Quaternaire. La baisse du niveau des mers, due à l’accumulation des glaces sur le Gondwana, aurait aussi été inférieure. Elle est évaluée à moins de 100 m, mais les plateaux continentaux étaient plus développés que durant le Quaternaire. C’était les parties immergées des continents, là où la vie abondait. Elle a donc été victime du rétrécissement et de la perte d’habitats, mais peut-être aussi d’un excès d’oxygénation due au changement des courants océaniques. La faune subsistante est dite à Hirnantia. Elle est caractérisée par des brachiopodes (Hirnantia, Eotropheodonta et Plectothyrella) et des trilobites. Les photos montrent des fossiles de brachiopodes d’Afrique du Nord. Le deuxième épisode d’extinction aurait été causé par la fin de la glaciation. D’abondants dépôts de black shales (schistes noirs) prouvent que des conditions anoxiques se sont installées. Ce sont des argiles mêlées à de la matière organique non oxydée et de la pyrite, liée à la décomposition de celle-ci. De tels dépôts sont susceptibles d’engendrer des hydrocarbures.
Durant l’Ordovicien, les températures de surface des mers ont chuté de manière très importante. Cette chute s’est accélérée durant l’Ashgill. Elle s’explique par la baisse de la teneur en dioxyde de carbone atmosphérique, qui devait être très élevée durant le Trémadocien. Selon les estimations, elle était comprise entre 14 et 22 fois la teneur actuelle (en 2012). Pour qu’une calotte glaciaire se forme sur le Gondwana, il a fallu qu’elle tombe à 8 fois le niveau actuel. L’altération des roches de la chaîne taconienne, soumise à de fortes pluies, a pu être un puits de CO₂, selon une théorie encore défendue en 2019 par Francis McDonald et ses collaborateurs.
En 2012, Timothy Lenton estimait que ce n’était pas suffisant. Pour lui et son équipe, l’altération des continents par la végétation a également joué un rôle : c’est durant l’Ordovicien qu’elle a commencé à gagner les terres émergées, jusqu’alors totalement dénudées. Des fossiles ressemblant à des spores d’hépatiques, de petites plantes non vasculaires, ont été datés à 473 Ma. Un tapis végétal fossilisé remonte aux 20 millions d’années suivantes. Grâce à une expérience effectuée avec une mousse moderne, Lenton a montré que ce tapis végétal démultiplie l’altération des roches, surtout des andésites, et le captage du CO2. Ainsi, la vie elle-même aurait contribué à cette extinction de masse.
Selon une hypothèse émise en 2004 par cinq scientifiques américains, la glaciation aurait été provoquée par une explosion de rayons gamma, qui aurait détruit la couche d’ozone de la stratosphère et provoqué l’irradiation des mers par les rayons ultraviolets. D’après eux, un tel phénomène, probablement causé par l’explosion d’une supernova, doit se produire au moins deux fois par milliard d’années. Mais cette hypothèse mal étayée n’a pas été retenue par la communauté scientifique. Elle est connue du grand public parce qu’elle a été reprise par la mini-série documentaire Animal Armageddon en 2009. Il fallait donner un caractère soudain et spectaculaire à cette extinction.












Leave a Reply
Vous devez vous connecter pour publier un commentaire.