


Un vaisseau extraterrestre qui aurait survolé notre planète il y a 3,5 milliards d’années n’y aurait vu aucune trace de vie. Elle était pourtant présente partout où l’eau était disponible, sur les continents comme dans les océans, car elle avait colonisé tous les milieux, les eaux douces comme les eaux salées. Elle s’installait dans des caldeiras de volcans et dans des sources chaudes, où les températures élevées ne la gênaient pas. L’oxygène, émis par les organismes photosynthétiques, n’était pas encore présent dans l’atmosphère, mais la vie pouvait se protéger des rayons ultraviolets, qu’aucune couche à ozone n’arrêtait. La Terre existait pourtant depuis plus d’un milliard d’années et de l’eau liquide était présente à sa surface depuis 900 millions d’années. Les chutes de météorites, abondantes en ces âges reculés, ne l’avaient pas éliminée. En s’approchant des lacs et des fleuves, le vaisseau aurait vu d’étranges rochers, les stromatolites. Il aurait fallu les analyser de près pour s’apercevoir qu’ils avaient été bâtis par des colonies de microbes : des cellules procaryotes, dépourvues de noyau. Il existait probablement déjà une distinction entre bactéries et archées.
Les ceintures de roches vertes de la Terre archéenne
À nous aussi, cette planète sur laquelle nous n’aurions pas pu respirer, faute d’oxygène, nous aurait paru exotique. Le Soleil était moins lumineux que maintenant. La Lune gravitait autour d’elle à une distance moindre et des effusions de lave se produisaient dans ses bassins. Cela causait sur Terre des marées beaucoup plus amples que maintenant. Comme la Terre tournait plus vite sur elle-même, les vents étaient plus puissants. Les océans, qui recouvraient presque toute la planète, étaient colorés en vert par des ions ferreux provenant des sources hydrothermales et des éruptions volcaniques. Ils comportaient certainement d’immenses plateaux basaltiques dus à des superpanaches mantelliques. Des noyaux de continents composés de granitoïdes, appelés des cratons, ont subsisté jusqu’à maintenant. À côté d’une tectonique des plaques primitive, avec des mouvements horizontaux de croûte et de la subduction, il existait des mouvements verticaux de croûte continentale (la sagduction).

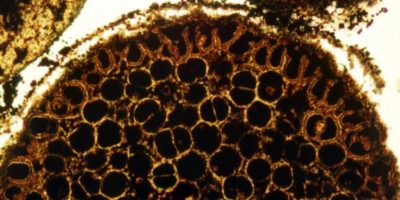
Des ceintures de roches vertes se formaient sur les continents, le plus souvent dans des zones immergées, grâce à d’abondantes coulées de laves basaltiques et komatiitiques. Les komatiites sont des roches volcaniques très pauvres en silice, que la Terre ne produit plus. Sur ces coulées, se déposaient des roches sédimentaires, dont les formations ferrifères rubanées (des minces couches alternées de silice microcristalline appelée du chert et de minéraux riches en fer), puis des argiles, des grès et des conglomérats. Ces derniers sont des accumulations de pierres provenant de l’érosion de reliefs, éventuellement emportées par des torrents. La ceinture de roches vertes de Barberton, sur le craton du Kaapval en Afrique du Sud, a des conglomérats datés à 3,22 Ga (milliards d’années), ce qui prouve l’existence de reliefs. Ses komatiites atteignent en revanche 3,48 Ga.
Les premières traces de vie doivent être recherchées dans ces sédiments. Des bactéries peuvent parfaitement être fossilisées et repérées au microscope. Elles deviennent des microfossiles. Le troisième éon de la Terre, le Protérozoïque (de 2,5 à 0,541 Ga), a livré des microfossiles parfaitement identifiés. Le chert de Gunflint, au Minnesota et dans l’Ontario, daté à 1,88 Ga, recèle les vestiges d’une véritable faune microscopique où se trouvaient certainement des eucaryotes – des cellules à noyau. Mais pour le deuxième éon de la Terre, l’Archéen (de 4 à 2,5 Ga), la tâche est beaucoup plus difficile, puisque les roches sont plus anciennes et souvent métamorphisées.
Les stromatolites
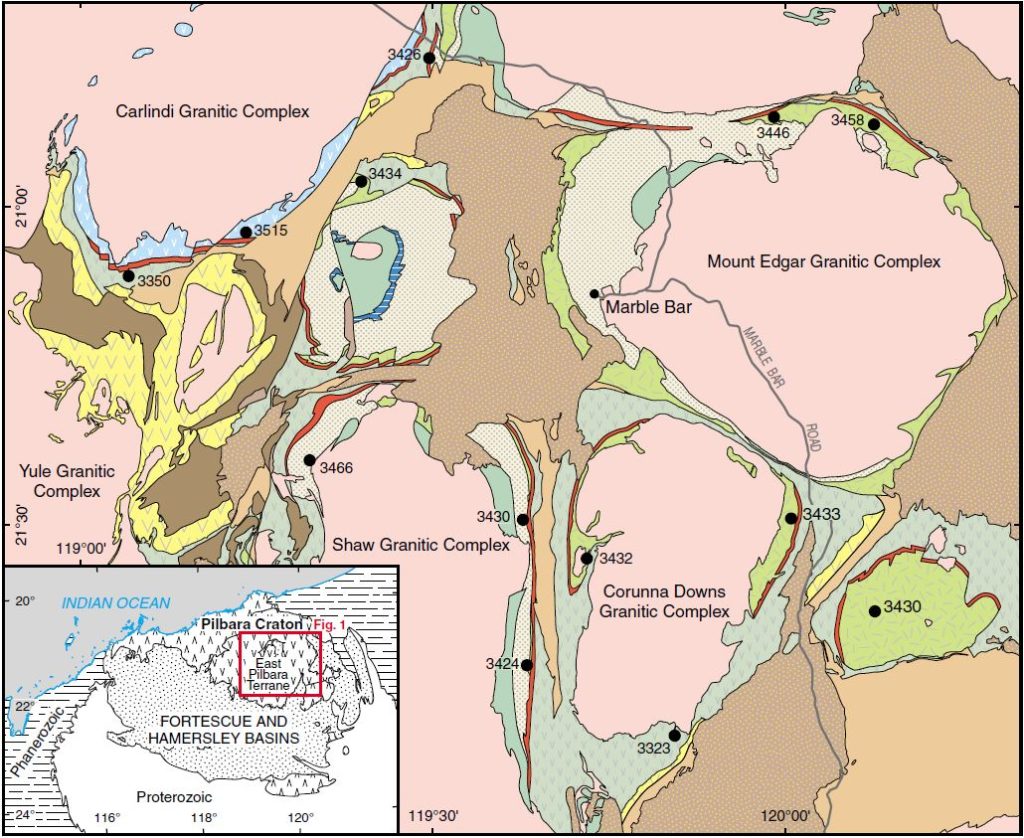
Un chercheur du Laboratoire d’Océanologie et de Géosciences de l’Université de Lille, Kevin Lepot, a fait le point sur ce problème dans un article publié en juillet 2020. Les stromatolites (ou stromatolithes) jouent un rôle primordial, mais ils peuvent être difficiles à reconnaître. Ce sont des structures laminées en forme de colonne, de cône ou de dôme, de composition carbonatée ou siliceuse. Il ne s’agit pas vraiment de fossiles, mais d’édifices laissés par des colonies microbiennes. Les stromatolites archéens contiennent rarement des microfossiles alors que ceux du Protérozoïque en sont parfois riches. En 2016, l’équipe d’Allen Nutman a annoncé la découverte de stromatolites dans la ceinture de roches vertes (également appelée la ceinture supracrustale) d’Isua, sur la côte nord-ouest du Groenland. Comme ils sont datés à 3,70 Ga, ils sont de 220 millions d’années plus anciens que les stromatolites de la formation Dresser sur le craton de Pilbara en Australie. Ceux-ci ont 3,48 Ga et leur origine biologique est certaine. L’identification des structures observées à Isua a été contestée par Mike Zawaski en 2020. Pour ce chercheur, elles sont d’origine métamorphique : ce sont des couches rigides de quartz et de dolomite (un carbonate), entourées de couches plus ductiles de schistes, qui ont été étirées et brisées. Ce phénomène est appelé un boudinage.

Les stromatolites de la formation de Dresser, appartenant au groupe de Warrawoona, ont été découverts dans les années 1980, mais leur identification a également été controversée, à cause de l’état d’altération de ces roches. Un examen des stromatolites présenté en 2019 a révélé qu’ils contiennent une grande part de pyrite FeS2 et qu’elle a des pores nanoscopiques, dans lesquels de la matière organique azotée a été repérée. Le carbone analysé a la composition isotopique du carbone organique. Les scientifiques ont également trouvé des filaments et de tresses caractéristiques des films microbiens. Par ailleurs, l’analyse des quatre isotopes stables du soufre a montré que ces micro-organismes effectuaient la réduction des sulfates SO42–, réaction qui libère du sulfure d’hydrogène H2S. Autrement dit, ils « respiraient » des sulfates pour obtenir de l’énergie. Ce métabolisme est attesté plus récemment, comme avec les stromatolites du groupe de Nsuze datés à 2,97 Ga. Ces sédiments font partie du supergroupe de Pongola en Afrique du Sud, et ils se sont déposés dans un milieu côtier soumis aux marées où la présence d’oxygène a été supposée.

Comme on le sait depuis 2015, grâce à Martin van Kranendonk, Kevin Lepot et d’autres scientifiques, la formation de Dresser s’est déposée dans la caldeira d’un volcan où des fluides hydrothermaux circulaient. Les stromatolites appartiennent à la couche inférieure, de 4 à 60 mètres d’épaisseur. Elle est composée de chert, de grès et de carbonates. L’activité hydrothermale y a laissé un réseau dense de veines de chert et de baryte (sulfate de baryum). Des roches édifiées par des geysers ont poussé M. van Kranendnok et David Deamer à chercher l’origine de la vie dans les sources chaudes et les mares volcaniques. Les sources hydrothermales océaniques restent cependant des berceaux plus probables pour la vie, mais elles n’ont pas été conservées. Comme il a été dit, les ceintures de roches vertes sont toutes continentales.

De même que la formation de Dresser, la formation de Strelley Pool fait partie des ceintures de roches vertes du craton de Pilbara (celle de Strelley), mais elle est plus récente. Elle a été datée entre 3,42 et 3,35 Ga. Ses stromatolites l’ont rendue célèbre, mais pour Kevin Lepot, seules les structures siliceuses columnaires sont authentiques. Elles présentent des ressemblances remarquables avec les stromatolites de Gunflint, riches en microfossiles, et même avec ceux du parc de Yellowstone. Ces derniers font partie des rares stromatolites qui soient toujours « vivants » aujourd’hui. Dans les stromatolites de Strelley Pool, des coagulations de particules carbonées sont visibles, mais ce ne sont probablement pas des microfossiles car elles sont trop petites ; elles sont submicrométriques. Ces stromatolites ont formé de véritables récifs. Les environnements dans lesquels les sédiments de Strelley Pool se sont déposés étaient des mers de faible profondeur, des plages, des lacs, des sebkhas (des lagunes soumises à une évaporation intense), des fleuves et des estuaires.

Les stromatolites de la formation de Tumbiana se trouvent également sur le craton de Pilbara, dans sa marge sud. Ils appartiennent au groupe de Fortescue, daté de 2,77 à 2,63 Ga, dont l’environnement était lacustre et fluvial. C’était une période de rift et le volcanisme était intense. Il a laissé d’épaisses coulées de basalte et des couches de cendres. Les stromatolites ont été édifiés dans un lac géant il y a 2,72 Ga. Très bien préservés, ils sont principalement composés de lamines de calcite microcristalline. La présence de globules sulfurisés de matière organique dans les lamines de calcite suggère que la réduction des sulfates par les microbes pourrait être un facteur important de cristallisation de calcite (organo-minéralisation) comme dans les stromatolites actuels. La réduction des ions ferriques Fe3+ était peut-être aussi à l’œuvre. Les stromatolites de Tumbiana contiennent d’abondantes structures appelées des fenestrae, qui pourraient être des reliques de bulles d’oxygène produites par la photosynthèse ou d’autres gaz. Ces structures sont également observées dans les stromatolites du supergroupe volcanique et sédimentaire de Ventersdorp (de 2,72 à 2,63 Ga) en Afrique du Sud. Ils ont une morphologie semblable à celle des stromatolites siliceux de Yellowstone. Les stromatolites du supergroupe de Pongola dans le même pays, âgés de 2,98 Ga auraient aussi des fenestrae résultant du piégeage d’oxygène dans des tapis microbiens.
La vie à l’origine de l’oxygène et du méthane dans l’atmosphère
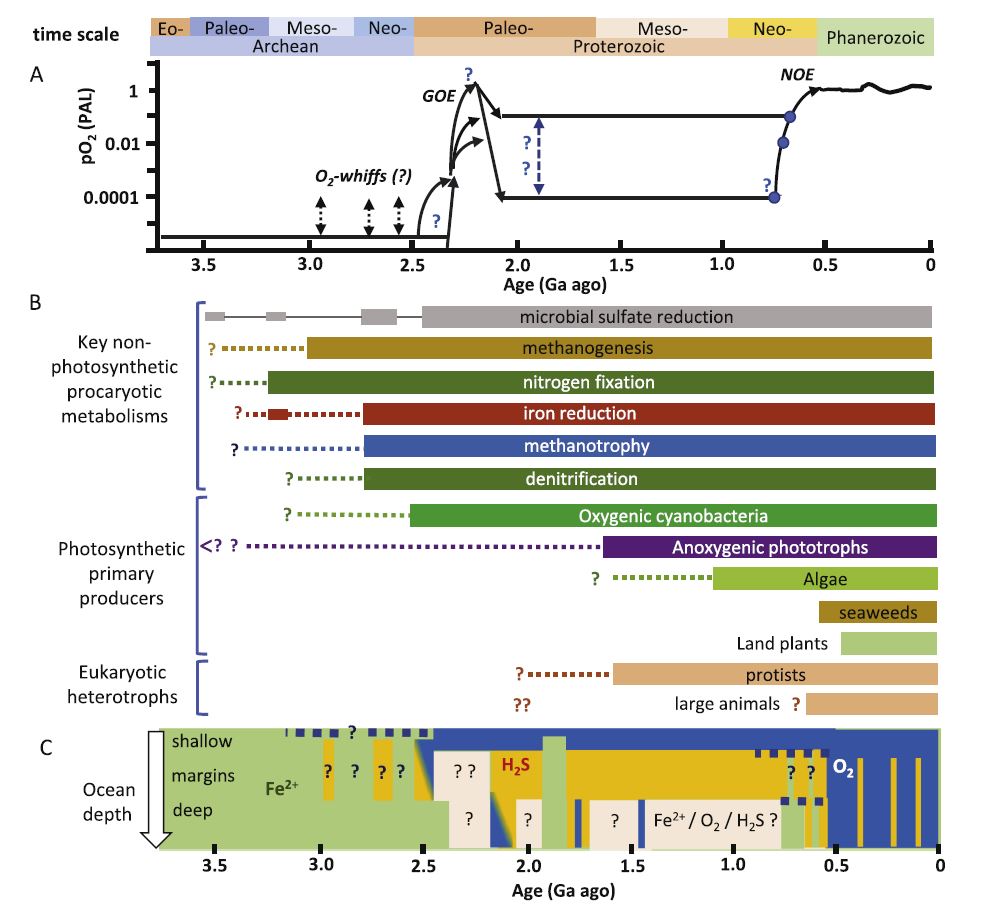
La photosynthèse oxygénique a très probablement existé durant l’Archéen, Il semble qu’elle ait commencé entre 3,2 et 2,8 Ga, durant une période appelée le Mésoarchéen. Pour la déceler, les chercheurs utilisent de plus en plus les isotopes stables du chrome. Cet élément est à 83,78 % composé de chrome 52 et à 9,50 % de chrome 53. Il possède aussi deux états naturels d’oxydation : le chrome (VI) soluble (CrO42– ou HCrO4–), qui est toxique, et les composés du chrome (III) insolubles. La réduction du chrome (VI) en chrome (III) se fait avec un fractionnement isotopique : la proportion 53Cr/52Cr varie. Cela fournit un moyen de connaître la concentration en oxygène dans des sédiments marins, tels que les formations ferrifères rubanées et les black shales (des argiles enrichies en matière organique). Les premières résultent de l’oxydation d’ions ferreux et de la précipitation de silice dans l’eau. Sur la base des isotopes du bore, la concentration en oxygène atmosphérique devait être égale à 0,03 % de la concentration actuelle il y a 3 Ga. Ce gaz participait à l’altération des roches. Le chrome (III) était oxydé dans les sols et du chrome (VI), isotopiquement lourd, était transporté par les rivières et les nappes phréatiques jusque dans les océans, où les ions ferreux en solution le réduisaient en chrome (III) et entraînaient sa précipitation.
Voir Une oasis d’oxygène dans un environnement côtier il y a 2,8 milliards d’années
Quand de l’oxygène était émis par des organismes photosynthétiques dans l’eau de mer, riche en ions ferreux, il entraînait la précipitation d’oxyhydroxydes de fer, une composante des formations ferrifères rubanées. La ceinture de roches vertes d’Iusa comporte de telles formations datées à 3,77 Ga, mais elles n’indiquent pas forcément l’existence de la photosynthèse oxygénique. C’est plus probablement la photoferrotrophie qui les a produites : une photosynthèse reposant sur l’oxydation des ions ferreux. Elle ne libère pas d’oxygène. Les formations ferrifères rubanées d’Isua indiquent tout de même l’existence de la vie durant le Paléoarchéen (de 3,6 à 3,2 G), alors que des stromatolites n’ont pas pu être identifiés au Groenland. Il existait des organismes qui « respiraient » du fer.

La libération de méthane par les organismes vivants a pu influer sur les climats de la Terre, car c’est un puissant gaz à effet de serre. Pour savoir si elle s’est produite durant l’Archéen, on examine les résidus de matière organique, notamment des kérogènes, qui sont susceptibles de libérer des hydrocarbures. Si leur carbone a été très appauvri en carbone 13 (l’isotope lourd de cet élément), c’est un indice de méthanogenèse. Elle est attestée durant la deuxième moitié du Mésoarchéen, de 3,0 à 2,8 Ga. Les organismes producteurs de méthane vivaient dans les lacs plutôt que dans la mer. Actuellement, ce sont des archées, mais les cyanobactéries sont également capables d’en produire. Les stromatolites de la formation de Dresser, influencés par l’activité hydrothermale, ont peut-être également libéré du méthane il y a 3,48 Ga.
De la vie dès l’Hadéen ?
Le stade ultime d’évolution du kérogène est le graphite : c’est du carbone pur. Sa composition isotopique pourrait indiquer une origine biologique, le carbone organique étant appauvri en 13C par rapport au carbone originaire du manteau terrestre. Il existe du graphite dans la ceinture supracrustale d’Isua, dans les roches de l’île d’Akilia, qui ont plus de 3,83 Ga, dans celles de la ceinture de roches vertes de Nuvvuagittuq au Québec, qui ont plus de 3,75 Ga, et dans celles du bloc de Saglek au nord du Labrador, qui ont environ 3,95 Ga. Prouver que ces graphites sont d’origine biologique augmenterait donc l’ancienneté des premières traces de vie. Le problème est que ces roches ont toutes subi un métamorphisme intense, du faciès des amphibolites à celui des granulites. Du graphite peut se former par décomposition thermique de carbonate de fer lors d’un tel métamorphisme. Néanmoins, une équipe japonaise dirigée par Takayuki Tashiro a pu démontrer en 2017 que du graphite du bloc de Saglek, contenu dans des sédiments métamorphisés, est d’origine biologique. Son δ13C est égal à – 28 ‰, alors que celui du carbone d’origine mantellique vaut – 5,5 ‰ et celui des carbonates marins est proche de 0 ‰. Cela signifie qu’il est très appauvri en carbone 13.
Il est possible de remonter encore plus loin dans le temps, jusqu’au premier éon de la Terre, l’Hadéen. Il va de 4 Ga jusqu’à la formation de notre planète il y a 4,57 Ga. Aucune roche hadéeenne n’a été conservée, mais il en subsiste des cristaux de zircon dont l’âge va jusqu’à 4,4 Ga. Leur présence implique celle d’eau liquide, dans laquelle la vie a pu apparaître. En 2015, l’équipe d’Elizabeth Bell a annoncé la découverte d’un zircon contenant une inclusion de graphite dont le δ13C est égal à environ – 24 ‰. On peut envisager une origine biologique, mais les précautions prises avec les métasédiments du bloc de Saglek ne peuvent pas être prises avec ce minuscule et unique cristal.
Tapis microbiens et microfossiles
Les cherts de Josefsdal âgés de 3,33 Ga, dans la ceinture de roches vertes de Barberton, contiennent un tapis microbien fossilisé identifié par l’équipe de Frances Westall. Il se situait dans un environnement côtier volcanique. Il n’était pas lié à des stromatolites et se développait grâce à des fluides hydrothermaux qui lui apportaient des nutriments, mais qui ont aussi permis une fossilisation rapide, car ils étaient riches en silice dissoute. Ces micro-organismes étaient thermophiles : ils appréciaient la chaleur.
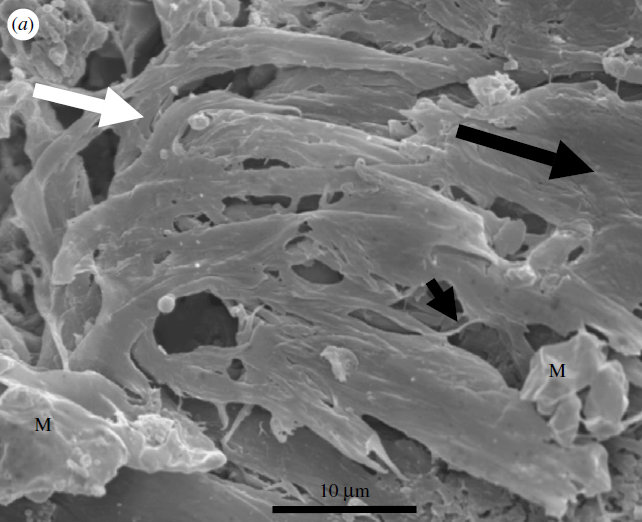
Des tapis microbiens d’un autre type ont été conservés. Des argiles ou d’autres sédiments fins s’y sont agglomérés, formant des structures caractéristiques de quelques millimètres de long qu’on appelle des MISS en anglais (Microbially Induced Sedimentary Structures). Les plus anciennes, âgées de 3,22 Ga, se trouvent dans le groupe de Moodies, qui fait partie de la ceinture de roches vertes de Barberton. Il est composé de sédiments détritiques déposés dans des milieux fluviatiles et marins peu profonds : des argiles, de la siltite (grès à grains très fins) et des grès surmontant des conglomérats. L’analyse des isotopes du carbone dans l’environnement terrestre indique l’existence d’une photosynthèse oxygénique ou anoxygénique utilisant le cycle de Calvin, peut-être couplée à la fixation de nitrates. Dans l’environnement marin, un appauvrissement accentué en carbone 13 a pu enregistrer de la sulfato-réduction microbienne, de la méthanogenèse ou de l’acétogenèse. Dans le dernier cas, de l’acétate est produit par fermentation. Des tapis microbiens enveloppant des grains détritiques âgés de 3,42 Ga pourraient se trouver dans la même ceinture de roches vertes, mais ce ne sont pas des MISS. Ils appartiennent au chert de Buck Reef. Des MISS potentielles existent également dans la formation de Dresser.

Les structures sphériques microscopiques sont de bonnes candidates pour être des microfossiles, mais elles peuvent également être d’origine purement abiotique, les minéraux sphériques étant fréquents. En 2010, l’équipe d’Emmanuelle Javaux a présenté une population de micro-organismes sphéroïdaux étonnamment grands, dont le diamètre atteint 300 µm, provenant du groupe de Moodies. Leur identification est certaine. Ils cohabitaient avec des tapis microbiens dans un milieu alluvial ou côtier.
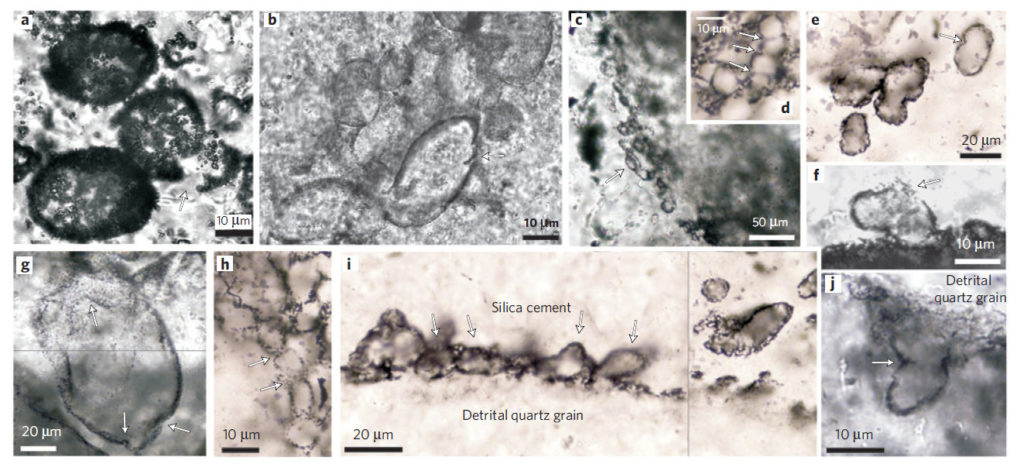
En 2011, David Wacey et Martin Brasier ont présenté des microfossiles plus anciens, de la formation australienne de Strelley Pool. À cet endroit, des roches volcaniques ont été datées à 3,51 Ga. Elles ont été érodées, puis la mer les a recouvertes et a constitué l’un des plus anciens rivages connus. Il y a environ 3,4 Ga, du sable produit par l’érosion de terres émergées, transformé plus tard en grès, s’est déposé dans un estuaire ou sur un littoral, sous une faible épaisseur d’eau. Les microfossiles sont situés à la base de cette formation, dans des grès noirs. Ils adhèrent parfois à des grains de sable, qui sont en quartz ou en pyrite. Aujourd’hui, on ne peut pas trouver de pyrite sur les plages, puisqu’elle est rapidement oxydée. Au milieu du Paléoarchéen, il n’y avait pas encore d’oxygène. La faune microbienne qui a vécu dans ce sable était variée : les microfossiles sont des sphéroïdes, des ellipsoïdes et des tubes cylindriques creux. Les dimensions vont de 2 à 25 µm. Des cristaux de pyrite micrométriques associés à ces fossiles sont des sous-produits de leur métabolisme : ces organismes effectuaient la sulfato-réduction. Les stromatolites de Strelley Pool sont postérieurs à ces communautés microbiennes.
Il existe d’autres microstructures susceptibles d’être des microfossiles, mais ceux du groupe de Moodies et de Strelley Pool sont parmi les plus spectaculaires.
****************************************************************************************
Kevin Lepot, Signatures of early microbial life from the Archean (4 to 2.5 Ga) eon, Earth-Science Reviews 209, 2020.
https://www.sciencedirect.com/science/article/pii/S0012825220303421












Leave a Reply
Vous devez vous connecter pour publier un commentaire.