

La photosynthèse effectuée par les végétaux, à la surface des océans et des continents, est l’unique source d’oxygène sur Terre. Par « oxygène », j’entends les molécules de dioxygène, formées de deux atomes d’oxygène, qui constituent un gaz. Les atomes d’oxygène sont également présents dans l’eau et dans toutes les roches, qu’elles soient silicatées ou carbonatées, mais ils sont liés à d’autres atomes d’une manière stable. La photosynthèse consiste à prélever du dioxyde de carbone dans les océans ou l’atmosphère, à le combiner à de l’eau pour former du glucose C6H12O6, ce qui libère de l’eau et de l’oxygène. Il ne s’agit ici que d’un bilan qui s’écrit :
6 CO2 + 6 H2O + énergie = C6H12O6 + 6 O2
De manière encore plus simplifiée, elle revient à scinder une molécule de CO2 pour en extraire C et libérer O2. La réaction inverse se produit également : c’est la respiration aérobie, dont le but est de libérer de l’énergie. Tous les végétaux la pratiquent, si bien que globalement, ils émettent autant de CO2 que d’O2. Il y a un cycle plus long à prendre en compte. Quand une plante est en période de croissance, elle fixe du carbone et émet de l’oxygène. Il se produit l’inverse à sa mort, car le carbone qu’elle contient est partiellement ou totalement oxydé lors de sa décomposition. La plante est alors une émettrice de CO2.

La biomasse totale des forêts est estimée à 1 650 Gt (milliards de tonnes), dont 1 025 Gt pour les seules forêts tropicales. C’est beaucoup plus que la biomasse océanique, estimée à 3,9 Gt et surtout concentrée sur les bordures continentales. En ajoutant les landes, les savanes, les steppes, les surfaces cultivées, etc., on arrive à une biomasse mondiale totale de 1 841 Gt. Les forêts en représentent 89,6 %. Ces chiffres ont été fournis par G. Ducreux en 2002. D’après un rapport du GIEC de 2013, il y a 2 300 Gt de carbone contenu dans les végétaux et les sols (sous forme d’humus notamment) et 3 Gt dans la biomasse océanique. Attention, il s’agit ici de la masse du carbone organique contenu dans la végétation et non de la masse de celle-ci. Les autres estimations donnent aux alentours de 550 Gt pour la masse du carbone végétal et de 1 500 Gt pour la masse du carbone contenu dans les sols.
On comprend l’importance des forêts tropicales, notamment de la forêt amazonienne. Cependant, elle n’est pas le « poumon de la planète » comme on le dit. Une forêt à l’équilibre ne fait pas varier le taux d’oxygène et de dioxyde de carbone dans l’atmosphère. En zone tempérée, une forêt émet de l’oxygène au printemps et du dioxyde de carbone en automne, si bien que le bilan annuel est nul. Mais en cas de sénescence, réduction ou destruction de la forêt, le flux de CO2 l’emporte sur celui d’O2. Voilà pourquoi toutes les forêts doivent être préservées, en particulier les forêts tropicales. Le déboisement fait pencher la balance en faveur du CO2 et contribue donc au réchauffement climatique. Néanmoins, cet effet est difficile à évaluer. Inversement, une augmentation des surfaces forestières entraîne un pompage du CO2 atmosphérique avec relâchement d’oxygène, le carbone étant fixé dans le bois et l’humus. C’est le meilleur puits de CO2. À cela, on peut ajouter que l’évapotranspiration augmente l’humidité des zones forestières : l’eau consommée par les plantes, grâce à leurs racines, est évacuée par les feuilles sous forme de vapeur d’eau. On estime par conséquent que la disparation d’une moitié de la forêt amazonienne pourrait entraîner une baisse de 20 à 30 % des précipitations sur la région.
C’est durant le Dévonien (de 416 à 359 Ma) que les continents ont été recouverts de forêts. Auparavant, il n’existait que de petites plantes inféodées aux cours d’eau, quasiment dépourvues de racines. L’arbre le plus ancien, Archaeopteris, est apparu durant le Dévonien. L’apparition et développement des forêts a dû entraîner une chute du taux de CO2. On estime qu’il est passé de 3 000 à 1 000 ppmv (parties par million en volume). Pour comparaison, il était de 280 ppmv avant l’ère industrielle. Cela aurait pu entraîner un important refroidissement climatique, mais la baisse de l’albédo de la Terre a permis à un climat chaud de se maintenir. L’albédo d’une surface est son pouvoir réfléchissant. Plus son albédo est faible, plus elle absorbe le rayonnement solaire et plus sa température augmente. Il est évident qu’une zone forestière absorbe plus de lumière qu’une zone de roches nues.
En toute logique, cette baisse du taux de dioxyde de carbone s’est accompagnée d’une hausse du taux d’oxygène, qui aurait jusqu’alors été d’environ 15 %. Il a continué à augmenter durant tout le Carbonifère (de 359 à 299 Ma), mais pas pour la même raison. C’est cette fois la fossilisation des forêts qui a joué. La lignine, composant majeur du bois, était alors mal décomposée, car les champignons supérieurs n’existaient pas encore. Les forêts qui poussaient dans des zones marécageuses au pied de l’imposante chaîne hercynienne étaient parfois inondées puis enfouies sous les sédiments, si bien qu’elles échappaient à l’oxydation. Cette matière organique s’est transformée en charbon. Grâce à ce processus, le taux d’oxygène a grimpé jusqu’à plus de 35 % au début du Permien (de 299 à 252 Ma).

Photo prise en 1970 par Werner Schellmann, Wikimedia Commons.
Il est important de comprendre que l’augmentation du taux d’oxygène est une conséquence de la sédimentation. Provenant de l’érosion des montagnes voisines, les sédiments s’accumulent sur les forêts mortes et empêchent l’oxydation de la matière végétale, laquelle peut se transformer en charbon. L’oxygène qui devrait être « consommé » par ce processus reste ainsi dans l’atmosphère. La raison de l’enrichissement de l’atmosphère en oxygène n’est donc pas la même au Carbonifère (de 359 à 299 Ma), époque à laquelle les veines de charbon ont commencé à se former, qu’au Dévonien. L’accumulation de matière organique mêlée à des argiles sur les planchers des mers conduit de manière comparable à la formation d’hydrocarbures dans des eaux anoxiques. L’oxygène qui a été libéré par les cyanobactéries et les algues durant leur vie reste dans l’atmosphère au lieu de contribuer à leur dégradation après leur mort.
Les sédiments dont je parle ici, qualifiés de détritiques, proviennent de l’érosion de terres émergées. Ce sont des sables, des silts et des argiles, les grains de silt ayant une taille intermédiaire entre ceux des sables et des argiles. Trois types de sédiments sont distingués : les sédiments marins, ceux des bassins houillers et les autres sédiments détritiques terrestres (red beds et arkoses). Les red beds, « lits rouges » en français, sont des sédiments détritiques colorés en rouge par les oxydes de fer, et les arkoses proviennent de sables riches en feldspaths. L’érosion qui les produit est capable de mettre à nu de la matière organique, telle que des strates comportant du charbon, qui avait jusque-là été préservée de l’oxydation. Elle est par conséquent responsable d’une baisse du taux d’oxygène.
En définitive, ce qui fait varier le taux d’oxygène, c’est la redistribution des sédiments entre les trois types qui ont été définis, mais la raison donnée plus haut de l’augmentation du taux à la fin du Carbonifère et au Permien (de 299 à 252 Ma) reste valable : elle est attribuée aux bassins houillers. La baisse du taux à la fin du Permien est probablement due à la disparition des marais causée par l’installation d’un climat sec sur la Pangée. La sédimentation se faisait alors plutôt sous forme de lits rouges exempts de carbone organique et de soufre. Je n’ai pas encore précisé qu’un autre mécanisme d’absorption d’oxygène est l’oxydation de la pyrite FeS2, minéral bien connu pour sa forme cubique et son éclat métallique, en partie formé lors de la décomposition de la matière organique. À la toute fin du Permien, une baisse généralisée du niveau des mers a mis à jour des veines de charbon qui ont alors été soumises à l’érosion et à l’altération, ce qui a contribué à la baisse du taux d’oxygène.
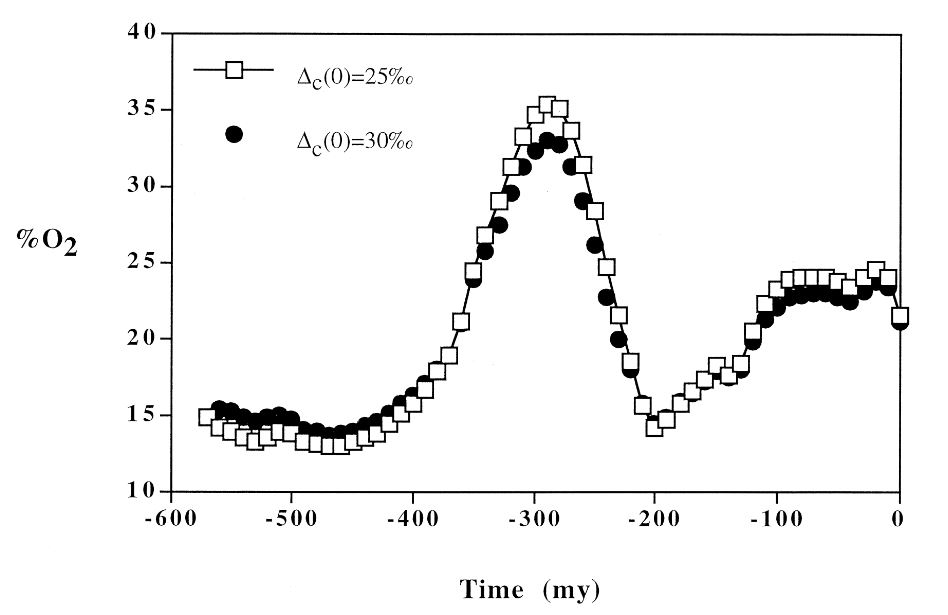
Robert A. Berner, Modeling atmospheric O2 over Phanerozoic time, Geochimica et Cosmochimica Acta, Vol. 65, No. 5, pp. 685– 694, 2001.












Leave a Reply
Vous devez vous connecter pour publier un commentaire.